


VALE A PENA EXPLORAR ESSE LINK
ESTUDO DIRIGIDO, RESUMOS, IMAGENS E INFOGRÁFICO
https://drive.google.com/file/d/1duDGx617acxbZ9ERerBOnhxb2z94LvTU/view?usp=sharing
O Imunoverso é um espaço para repensar nossa relação com o mundo microscópico. Por muito tempo, descrevemos a interação entre nosso corpo e os microrganismos usando metáforas de guerra, invasão e conquista — uma linguagem herdada de uma visão eurocêntrica de mundo focada em dominação. Mas a biologia nos conta uma história muito mais complexa e interconectada. Vamos explorar as infecções não como uma batalha, mas como um processo dinâmico de negociação evolutiva.
Nossa espécie, Homo sapiens, surgiu há cerca de 300.000 anos em um planeta que já era um ecossistema vibrante e plural. Nossos ancestrais não evoluíram em um mundo estéril; eles emergiram imersos em um oceano de vida, desde as bactérias do solo até os vírus carregados por outros animais. Desde o primeiro momento, a coexistência não foi uma opção, mas a própria condição da nossa existência. O parasitismo, portanto, não é uma anomalia ou um ataque externo, mas uma das mais antigas e fundamentais formas de interação ecológica.
Quando um patógeno encontra um novo hospedeiro, o que muitas vezes vemos não é uma “guerra”, mas uma Crise Adaptativa de Rejeição Mútua. É um estado de profundo desequilíbrio, uma incompatibilidade biológica onde a replicação do microrganismo e a resposta defensiva do hospedeiro se retroalimentam de forma insustentável. O sistema imunológico, em sua tentativa de rejeitar o organismo estranho, pode gerar inflamações e febres que danificam o próprio corpo. Do outro lado, a multiplicação do patógeno rejeita homeostase do hospedeiro, alterando seu funcionamento para seu próprio benefício.
Essa crise, no entanto, é um estado evolutivamente instável. O objetivo final de qualquer organismo não é a destruição, mas a perpetuação. Um patógeno que elimina rapidamente seu hospedeiro muitas vezes encontra um beco sem saída evolutivo. Por isso, a seleção natural frequentemente favorece caminhos que levam para fora da crise. Ao longo de gerações, as linhagens de patógenos que moderam sua virulência e as linhagens de hospedeiros que desenvolvem uma resistência mais calibrada são as que tendem a prosperar. O conflito agudo se transforma gradualmente em uma coexistência tensa, uma negociação contínua onde o dano é minimizado para garantir a sobrevivência de ambos.
Esse espectro de interações é a verdadeira face da coevolução. Ele vai desde o desequilíbrio violento de uma nova epidemia até a convivência estável de longo prazo, como a dos vírus do herpes que habitam nosso corpo por toda a vida. Em casos extremos, essa negociação pode levar à cooperação total, como aconteceu com as antigas bactérias que, um dia, se tornaram as mitocôndrias dentro de nossas células, a base da nossa própria vida complexa. Entender o parasitismo é entender que a vida não evolui através de conquistas, mas através de interações, adaptações e da incessante busca por equilíbrio em um mundo compartilhado.
1. Organização Corporal e Holobionte
A compreensão contemporânea do corpo humano exige o abandono da visão clássica do indivíduo como uma fortaleza biológica isolada, definida por fronteiras genéticas rígidas e protegida por um exército imunológico vigilante contra o “estrangeiro”. Em vez disso, a imunologia moderna, ao dialogar com perspectivas decolonais, propõe a noção de holobionte, um conceito que redefine o ser humano como um ecossistema integrado, composto pela hospedeiro e por sua vasta e essencial comunidade de microrganismos simbiontes. Essa mudança de paradigma requer uma profunda desconstrução das metáforas bélicas de invasão e defesa que historicamente dominaram o pensamento científico ocidental, as quais frequentemente refletiam lógicas coloniais de exclusão e pureza. No contexto do holobionte, o sistema imunológico deixa de ser visto meramente como um mecanismo de destruição do que é “não-eu” e passa a ser compreendido como um órgão de percepção e mediação relacional, um diplomata molecular encarregado de gerir a convivência entre múltiplas espécies. Essa organização se fundamenta na simpoiese, ou o “fazer-se com”, onde a identidade biológica não é algo dado ou estático, mas um processo contínuo de criação compartilhada entre células humanas, bactérias, fungos e vírus. Através da simbiose, esses agentes não humanos participam ativamente da maturação das respostas imunes, da regulação metabólica e até do desenvolvimento neurológico, tornando a distinção entre “eu” e “outro” uma fronteira fluida e porosa. Assim, a imunologia decolonial nos convoca a perceber que a vida não prospera no isolamento, mas na interdependência radical, onde a saúde é o resultado de uma diplomacia biológica bem-sucedida que sustenta a harmonia dentro dessa assembleia multiespécie. O corpo, portanto, não é uma propriedade individual, mas um território de coexistência e coevolução, onde a imunidade atua como a linguagem que permite ao holobionte reconhecer, acolher e tecer a complexa rede de conexões que sustenta a integridade da vida em sua plenitude ecológica.
O corpo é um sistema complexo composto por diversas unidades funcionais. A organização biológica pode ser compreendida em múltiplos níveis:
Holobionte: Refere-se à entidade biológica composta por um hospedeiro e todos os seus microrganismos associados (microbiota). Essa interação é fundamental para a saúde e o funcionamento imunológico.
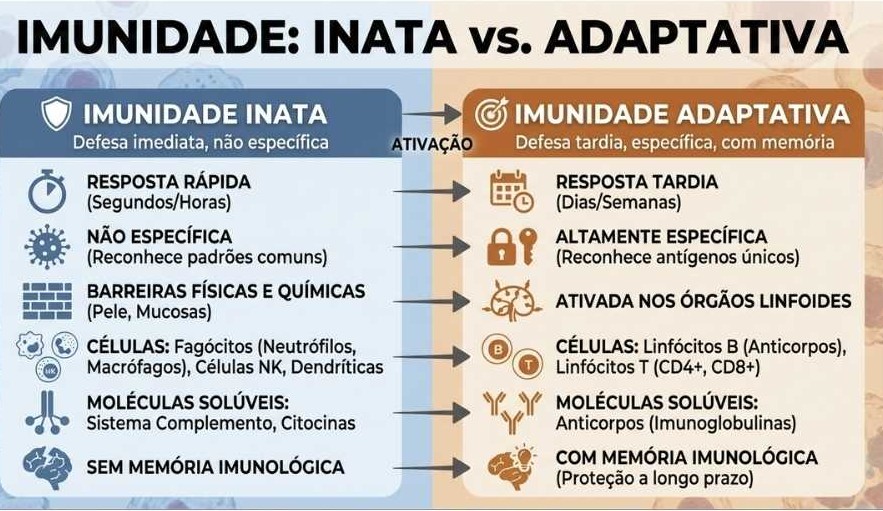
Tipos de Resposta Imune
Imunidade Inata: Primeira linha de defesa é formada por células e estruturas que agem de forma inespecífica, ou seja, independente de qual for o estímulo, a ação será sempre da mesma forma, agindo rapidamente. A ausência de especificidade não significa que essa resposta não possa ser ‘treinada’ a agir mais intensamente.
Imunidade Adaptativa: Essa imunidade ela é induzida a se maturar, ou seja a se adaptar à condição estressante, ela entra em ação tempos depois do estímulo e para que ocorra, células específicas ao estímulo, são selecionadas para responderem. Essa resposta é formada por Linfócitos T e B. A imunidade adaptativa pode ser celular e humoral.
Vamos falar bastante a palavra ‘antígeno’.
Um antígeno é qualquer ‘substância’ que o sistema imunológico consegue reconhecer e estabelecer uma resposta imunológica. Antígenos podem ser ‘próprios’ – que pertencem àquele holobionte (corpo). ‘não-próprios’ – moléculas externas ao ecossistema habitual desse holobionte. Em se tratando de condições nas quais esse holobionte reage de forma exagerada à moléculas externas que são normalmente inofensivas para demais holobiontes – condições de alergias, esse ‘antígeno’ é chamado alérgeno.
2. Organização Celular e Tecidual
A organização celular e tecidual do sistema imune no holobionte humano não se estrutura como uma muralha estática, mas como uma sofisticada rede de assembleias multiespécies distribuída em geografias de encontro e diálogo permanente. Sob a lente da simpoiese e da imunologia decolonial, os tecidos de barreira, como a mucosa intestinal, a pele e os pulmões, deixam de ser compreendidos como perímetros de isolamento para serem vistos como zonas de contato onde a arquitetura biológica é co-construída pela interação constante entre células do hospedeiro e comunidades microbianas. No trato gastrointestinal, por exemplo, o Tecido Linfoide Associado ao Intestino (GALT) opera como um centro nevrálgico de diplomacia molecular, onde a própria existência das Placas de Peyer e a maturação de folículos linfoides dependem umbilicalmente de sinais químicos emitidos pelos simbiontes. A nível celular, o conceito de exército de defesa é substituído por uma ecologia de mediadores: células dendríticas e macrófagos não atuam como meros carrascos do estranho, mas como tradutores biofísicos que amostram o ambiente e distinguem, através de um aprendizado relacional, entre o ruído da ameaça e a harmonia da cooperação. Essa organização é intrinsecamente descentralizada e plural, pois o sistema imune “estendido” incorpora os próprios microrganismos, que produzem bacteriocinas, ácidos graxos de cadeia curta e metabólitos que regulam o tônus inflamatório e a diferenciação de linfócitos T reguladores, fundamentais para a manutenção da paz tecidual. Assim, a imunidade não é uma propriedade privada do genoma humano, mas um fenômeno emergente de um “corpo-território” onde linfócitos intraepiteliais e células imunes inatas coabitam e coevoluem com vírus, fungos e bactérias em uma trama de interdependência radical. Ao decolonizar a visão tecidual, percebemos que a integridade do holobionte é sustentada por uma infraestrutura de hospitalidade biológica, onde a saúde não é a ausência do outro, mas a capacidade tecidual de sustentar uma simbiose dinâmica, transformando o que antes era chamado de “invasão” em um processo contínuo de fazer-se com o mundo através de uma biologia da partilha e da coexistência.
Células: As unidades básicas da vida, com funções especializadas. No contexto imunológico, diversas células atuam na defesa do organismo.
Tecidos: Agregados de células semelhantes que desempenham funções específicas (ex: tecido epitelial, tecido conjuntivo, tecido muscular, tecido nervoso).
Órgãos: Estruturas formadas por diferentes tipos de tecidos que cooperam para uma função particular (ex: coração, pulmão, baço).
3. Vascularização Sanguínea e Linfática
A circulação sanguínea e linfática, dentro da perspectiva do holobionte, deixa de ser compreendida como um sistema de encanamento para transporte de tropas e passa a ser vista como um território fluido de comunicação e interdependência que interliga os diversos nichos ecológicos do corpo-território. O sangue atua como o grande rio da diplomacia sistêmica, onde o fluxo constante de células imunes, nutrientes e, crucialmente, metabólitos derivados da microbiota, garante que nenhuma parte do ecossistema humano permaneça isolada. Através do sistema circulatório, sinais moleculares produzidos por simbiontes em locais distantes, como o intestino, viajam para influenciar a hematopoese na medula óssea e a prontidão imunológica em órgãos como o cérebro e os pulmões, evidenciando que a resposta imune não é um evento local, mas um processo de ressonância holística. De forma complementar, a circulação linfática opera como uma rede de escuta e acolhimento das particularidades teciduais, onde o líquido intersticial, carregado de informações sobre o estado de equilíbrio da vizinhança celular, é gentilmente conduzido aos linfonodos. Estes centros não devem ser interpretados sob a lógica decolonial como postos de vigilância militar ou alfandegas de fronteira, mas sim como câmaras de ressonância e assembleias multiespécies onde a apresentação de antígenos ocorre como um ato de tradução cultural entre diferentes formas de vida. Nos linfonodos, a linfa traz consigo não apenas fragmentos do que é “estranho”, mas as notícias químicas da convivência simbiótica, permitindo que os linfócitos sejam educados não para a exclusão sistemática, mas para o reconhecimento da complexidade. A migração celular — o trânsito constante de linfócitos T e B entre o sangue e a linfa — representa a circulação da memória e da experiência coletiva do holobionte, garantindo que o aprendizado obtido em uma zona de contato seja compartilhado com o todo. Esse dinamismo circulatório é a expressão máxima da simpoiese, onde o movimento do fluido é o que permite ao organismo “fazer-se com” o seu entorno, transformando a resposta imune em um fluxo contínuo de hospitalidade e ajuste mútuo. Assim, o coração e os vasos linfáticos não são apenas bombas e dutos, mas os facilitadores de uma biopolítica da partilha, onde a integridade da vida é sustentada pela capacidade de circular, integrar e responder coletivamente aos desafios de um mundo em constante interrelação.
Sistema Sanguíneo: Rede de vasos (artérias, veias, capilares) que transporta sangue, oxigênio, nutrientes, hormônios e células imunes por todo o corpo.
Sistema Linfático: Rede de vasos (vasos linfáticos) e órgãos (linfonodos, baço, timo) que transporta linfa, um fluido contendo células imunes e produtos residuais. Desempenha um papel crucial na vigilância imunológica e na remoção de patógenos.
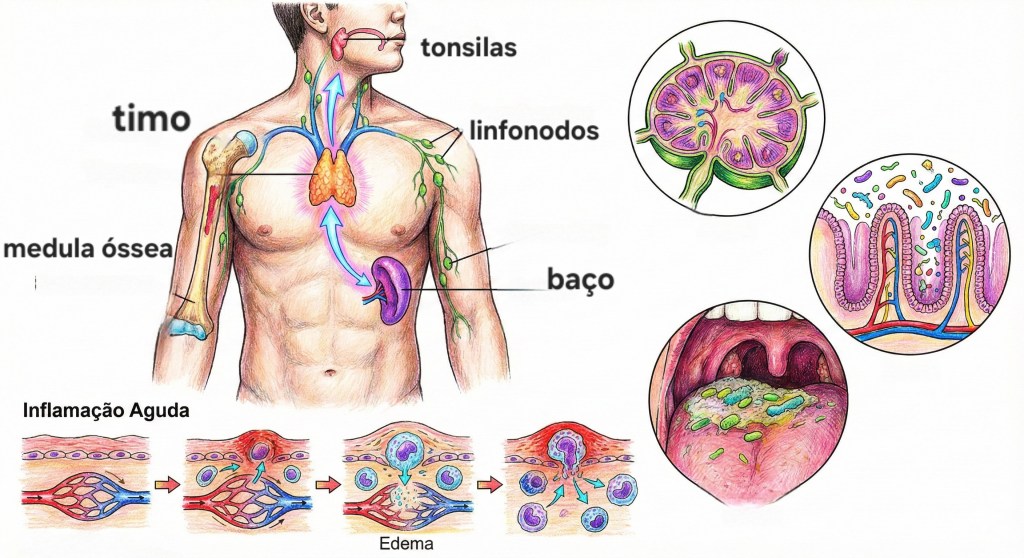
4. Órgãos e Tecidos Linfoides
A arquitetura dos órgãos e tecidos linfoides, sob a ótica da imunologia decolonial e do conceito de holobionte, deixa de ser interpretada como uma rede de quartéis e campos de treinamento para ser compreendida como uma geografia de maturação e espaços de assembleia onde se cultiva a arte da coexistência. Os órgãos linfoides primários, a medula óssea e o timo, funcionam como berços ontológicos onde a potencialidade da vida imunológica é gestada, mas não em um vácuo de pureza biológica. Na medula óssea, a gênese das linhagens celulares não é apenas um programa genético isolado do hospedeiro, mas um processo de “fazer-se com” (simpoiese) que responde aos sinais químicos e metabólicos que emanam de todo o holobionte, onde a presença de simbiontes distantes influencia o ritmo e a qualidade da produção celular. O timo, por sua vez, atua como uma escola de diplomacia e alteridade, onde a seleção dos linfócitos T não visa a criação de uma força de exclusão, mas o aprendizado de um reconhecimento relacional; é neste espaço que o organismo define os termos de sua hospitalidade, aprendendo a distinguir entre a agressão que rompe o equilíbrio e a alteridade que compõe a harmonia do ser. Uma vez formadas, essas células não se dirigem a postos de batalha, mas migram para os órgãos linfoides secundários — como os linfonodos, o baço e os tecidos linfoides associados às mucosas (MALT) —, que operam como verdadeiras praças públicas e casas de encontro multiespécies. Especialmente no MALT, a distinção entre o “eu” e o “outro” se dissolve em uma trama de interdependência radical, pois a própria organização tecidual desses locais é co-construída pela presença ativa da microbiota, que estimula a formação de centros germinativos e a diversificação de anticorpos. Nesses territórios de encontro, o baço filtra as notícias do fluxo sanguíneo enquanto os linfonodos processam os relatos da linfa, funcionando não como alfândegas de controle rígido, mas como centros de síntese onde o holobionte avalia e recalibra sua integridade ecológica. Assim, a organização linfoide não é uma estrutura de defesa de uma fortaleza, mas uma infraestrutura de comunicação e cuidado que permite ao organismo habitar a sua própria complexidade, transformando cada tecido em um espaço de negociação constante e cada órgão em um nó de uma vasta rede simbiótica que sustenta a vida como um fenômeno coletivo e territorial
Órgãos Linfoides Primários:
Medula Óssea: Local de produção de todas as células sanguíneas, incluindo as células imunes.
Timo: Local de maturação dos linfócitos T.
Órgãos Linfoides Secundários:
Linfonodos: Filtram a linfa e são locais de encontro entre antígenos e linfócitos.
Baço: Filtra o sangue e remove células sanguíneas velhas, além de ser um local de resposta imune contra patógenos sanguíneos.
Tecido Linfoide Associado à Mucosa (MALT): Inclui as amígdalas, adenoides, placas de Peyer (no intestino) e apêndice, protegendo as superfícies mucosas.
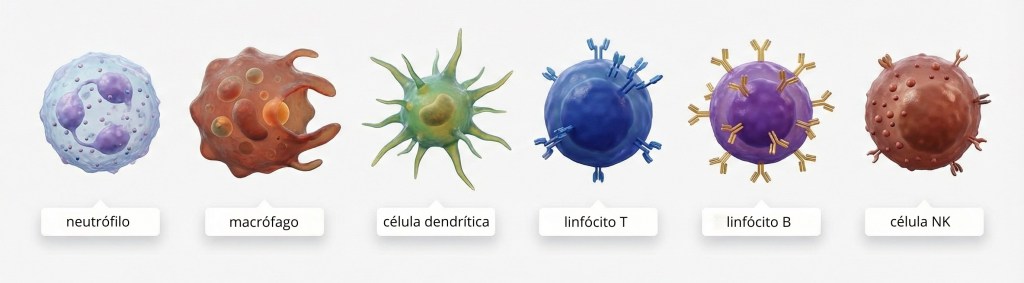
5. Células do Sistema Imune: Tipos, Origens e Funções

As células do sistema imune, em sua vasta diversidade de formas e propósitos, não emergem de um programa genético isolado e autorreferencial, mas de um processo de hematopoese que é, essencialmente, um ato de simpoiese ou “fazer-se com o mundo”. Originadas no território da medula óssea, as linhagens mieloide e linfoide não são meros soldados produzidos por uma fábrica biológica, mas agentes mediadores cuja maturação é sensível aos sussurros bioquímicos da microbiota e do ambiente. Sob a perspectiva decolonial, a origem dessas células deixa de ser vista como o nascimento de um exército de defesa para ser compreendida como a formação de uma comunidade de tradutores e cuidadores da integridade do holobionte. As células da linhagem mieloide, como os macrófagos e os neutrófilos, atuam como os jardineiros e sentinelas do equilíbrio tecidual, onde sua função primordial não é o extermínio cego do “outro”, mas a manutenção da harmonia através da fagocitose — um processo de incorporação e reciclagem que sustenta a renovação da vida. Os macrófagos, em particular, funcionam como historiadores locais dos tecidos, interpretando sinais de estresse e reparando danos, agindo mais como diplomatas que restauram a paz do que como combatentes em uma guerra perpétua. As células dendríticas, por sua vez, representam a ponte da inteligência relacional, capturando fragmentos do ambiente e da microbiota para apresentá-los nas assembleias dos órgãos linfoides, realizando uma tradução molecular que educa o organismo sobre quem são os parceiros da simbiose e quais são os sinais de ruptura da ordem ecológica. Na linhagem linfoide, os linfócitos T e B e as células NK manifestam a capacidade do holobionte de cultivar uma memória afetiva e específica das suas interações com o mundo. Em vez de operarem sob uma lógica de exclusão xenofóbica, essas células aprendem a reconhecer a alteridade através de processos de seleção que, numa visão decolonial, são entendidos como o desenvolvimento de uma hospitalidade crítica. Os linfócitos T reguladores exemplificam essa função ao atuar como os tecedores da tolerância, garantindo que a presença de microrganismos essenciais não dispare respostas agressivas, mas sim uma coexistência produtiva. As células B e seus anticorpos não são apenas projéteis de defesa, mas moléculas de adesão e reconhecimento que ajudam a organizar a arquitetura da microbiota nas mucosas, mediando quem permanece e como se distribui na vasta rede do holobionte. Assim, as funções celulares são ressignificadas: a imunidade não é uma arma contra o estrangeiro, mas a capacidade sensível de um corpo-território multiespécie de gerir suas próprias fronteiras porosas, transformando a vigilância em cuidado e a resposta imune em uma prática contínua de diplomacia biológica que sustenta a integridade da vida em comum.
O sistema imune é composto por diversos tipos de células, cada uma com funções especializadas:
Linhagem Mieloide:
Neutrófilos: Primeiras células a chegar ao local da infecção, fagocitam patógenos.
Eosinófilos: Envolvidos na defesa contra parasitas helmintos e reações alérgicas.
Mastócitos, Basófilos: Liberam histamina e outros mediadores da inflamação são células envolvidas, assim como os eosinófilos nas reações alérgicas e defesa contra parasitas helmintos.
Monócitos/Macrófagos: Fagocitam patógenos, apresentam antígenos e liberam citocinas. Os monócitos são encontrados circulando no sangue, quando migram para os tecidos se diferenciam em macrófagos e podem também se diferenciarem em células dendríticas. Os macrófagos apresentam antígenos para linfócitos T efetores (ativados anteriormente).
Células Dendríticas: Excelentes apresentadoras de antígenos, ativam linfócitos T virgens. São as células especializadas em levar o antígeno do local de sua presença aos órgãos linfoides secundários onde buscarão e farão interações com os linfócitos T específicos.
Linhagem Linfoide:
Linfócitos B: São células que reconhecem especificamente um antígeno, conseguem capturar o antígeno, processar e apresentar aos linfócitos T para a mediação da resposta imune humoral Timo dependente, podem se diferenciar em plasmócitos e produzem anticorpos.
Linfócitos T:
Linfócitos T Auxiliares (CD4+): Coordenam a resposta imune.
Linfócitos T Citotóxicos (CD8+): Matam células infectadas ou tumorais.
Linfócitos T Regulatórios (Tregs): Suprimem a resposta imune para evitar autoimunidade.
Células Natural Killer (NK): Matam células infectadas por vírus e células tumorais sem necessidade de ativação prévia, pertencem à imunidade inata e efetuam a morte da célula-alvo por meio de mecanismos similares aos ativados por linfócitos T CD8+.
6. Moléculas Secretadas: Citocinas, Quimiocinas e Sistema Complemento

As moléculas secretadas pelo sistema imune — citocinas, quimiocinas e as proteínas do sistema complemento — constituem a sofisticada gramática molecular que permite ao holobionte humano narrar sua existência e coordenar a vida em comum. Longe de serem meros “projéteis químicos” ou “alarmes” de uma guerra biológica, essas substâncias são os fios de uma trama simpoética, os mediadores de uma diplomacia biosemiótica que sustenta a comunicação entre as células do hospedeiro e a vasta comunidade de simbiontes. As citocinas operam como a voz do coletivo, uma sinfonia de sinais que modula a intensidade e a qualidade das relações teciduais; elas não apenas sinalizam o perigo, mas traduzem o estado de equilíbrio do ecossistema, definindo se o momento exige uma resposta de cuidado, de reparo ou de contenção. Sob uma perspectiva decolonial, a produção de citocinas não é vista como uma ordem hierárquica de comando e controle, mas como um diálogo polifônico onde os metabólitos da microbiota participam ativamente da “afinação” do tom inflamatório, ensinando ao corpo que a saúde não é o silêncio, mas uma ressonância harmoniosa entre diferentes formas de vida. As quimiocinas, por sua vez, funcionam como os cartógrafos da hospitalidade, estabelecendo gradientes químicos que orientam o trânsito das células imunes através das geografias do corpo-território. Elas não apenas “recrutam tropas”, mas convidam os agentes mediadores a habitarem as zonas de contato onde o diálogo com o outro se faz mais necessário, garantindo que o movimento celular seja um ato de encontro e não apenas de confronto. Entrelaçado a esse fluxo comunicativo, o sistema complemento emerge como uma rede ancestral de proteínas plasmáticas que, em vez de ser reduzida a uma cascata de destruição, é compreendida como um sistema de marcação e reconhecimento que facilita a integração e a limpeza do espaço comum. A opsonização, nesse contexto, torna-se um ato de “nomeação” molecular, permitindo que o holobionte identifique e processe componentes da sua própria rede, mantendo a fluidez das trocas metabólicas. Essas moléculas secretadas formam, portanto, uma ecologia de mensagens que ignora as fronteiras fixas entre o “eu” e o “outro”, revelando que a imunidade é um fenômeno de rede, onde a informação circula para sustentar a interdependência radical. Ao decolonizar o olhar sobre essas substâncias, percebemos que a vida no holobionte é tecida por essa química da partilha, na qual cada molécula é um gesto de manutenção de uma paz dinâmica e cada resposta imune é uma renegociação coletiva do compromisso de viver e fazer-se com o mundo.
Citocinas: Proteínas que atuam como mensageiros entre as células imunes, regulando a intensidade e a natureza da resposta imune (ex: interleucinas, interferons, fator de necrose tumoral).
Quimiocinas: Um tipo específico de citocina que atrai células imunes para locais de inflamação ou infecção.
Sistema Complemento: Um conjunto de proteínas plasmáticas que, uma vez ativadas, participam da eliminação de patógenos por meio de lise direta, opsonização e recrutamento de células imunes.
7. Receptores e Ligantes
A constelação de receptores e ligantes que define a dinâmica molecular do holobionte constitui a base sensorial de uma inteligência compartilhada, transcendendo a visão reducionista de mecanismos de “chave e fechadura” voltados para o controle de fronteiras. Nesta perspectiva simpoética, os Receptores de Reconhecimento de Padrão (PRRs), como os receptores Toll-like (TLRs) e NOD-like (NLRs), não atuam como dispositivos de detecção de inimigos, mas como órgãos de percepção da vizinhança que captam a “perfumaria molecular” da microbiota. Seus ligantes chamados de padrões moleculares associados a patógenos (PAMPs). Essa interlocução não busca o extermínio, mas a calibração da convivência, onde o reconhecimento de um ligante microbiano pode desencadear tanto um alerta de reparo quanto uma carícia bioquímica que fortalece a barreira epitelial. No coração dessa diplomacia molecular, o Complexo Principal de Histocompatibilidade (MHC) deixa de ser visto como um selo de identidade nacional ou um crachá de “eu” biológico para ser compreendido como um palco de oferendas, onde fragmentos da vida interna da célula — sejam eles derivados do metabolismo próprio ou de inquilinos intracelulares — são expostos para a apreciação coletiva. Os receptores de células T (TCR) e de células B (BCR), por sua vez, representam a capacidade do holobionte de gerar uma diversidade infinita de formas de encontro; sua geração através da recombinação somática não é um armamento preventivo, mas um exercício de poesia biológica que permite ao sistema imune estar pronto para reconhecer a alteridade em suas formas mais imprevistas. Os ligantes desses receptores, os antígenos, tornam-se assim pontes de reconhecimento que conectam a experiência individual à memória da espécie e à presença do outro. Além desses, os receptores de citocinas e quimiocinas atuam como os ouvidos da rede, captando os matizes das mensagens enviadas por outros agentes, enquanto os receptores de checkpoint imunológico, como o PD-1 e o CTLA-4, funcionam como moderadores da hospitalidade, garantindo que o entusiasmo do reconhecimento não se transforme em uma inflamação que agrida o território comum. Mesmo o sistema complemento, com seus ligantes e receptores de fragmentos de C3 e C5, é integrado nessa lógica como uma teia de marcação afetiva que facilita a limpeza e a organização dos espaços intersticiais. Ao decolonizar a compreensão desses processos, percebemos que o complexo de receptores e ligantes não é uma rede de espionagem, mas uma infraestrutura de escuta radical, onde a vida se reconhece, se comunica e se ajusta em um fluxo contínuo de simbiose, transformando o corpo em um território de hospitalidade consciente onde cada ligação molecular é um compromisso renovado com a integridade do coletivo multiespécie.
A comunicação entre as células imunes e o reconhecimento de patógenos são mediados por receptores e seus ligantes.
TCR (Receptor de Células T): Reconhece antígenos apresentados por moléculas MHC nas células apresentadoras de antígenos.
BCR (Receptor de Células B): Reconhece antígenos diretamente.
PRR (Receptores de Reconhecimento de Padrão): Reconhecem padrões moleculares associados a patógenos (PAMPs) e padrões moleculares associados a danos (DAMPs).
- PAMPs (Padrões Moleculares Associados a Patógenos): Moléculas conservadas encontradas em microrganismos (ex: LPS bacteriano, RNA viral).
- DAMPs (Padrões Moleculares Associados a Danos): Moléculas liberadas por células hospedeiras danificadas ou morrendo.
8. Opsonização e ADCC (Citotoxicidade Celular Dependente de Anticorpos)
Dentro da dinâmica de manutenção da harmonia do holobionte, os processos de opsonização e citotoxicidade celular dependente de anticorpos (ADCC) revelam-se não como táticas de extermínio sumário, mas como rituais de marcação e responsabilidade coletiva que garantem a integridade do corpo-território. A opsonização, tradicionalmente descrita como o ato de tornar um patógeno “saboroso” para a fagocitose, é aqui ressignificada, sob uma perspectiva decolonial, como um gesto de nomeação molecular e visibilidade. Através do depósito de anticorpos e proteínas do sistema complemento sobre a superfície de agentes que romperam o pacto da simbiose, o sistema imune cria um “manto de reconhecimento” que sinaliza a necessidade de uma intervenção cuidadosa. Este processo não visa meramente a destruição, mas a identificação de elementos que, por estarem em desequilíbrio ou por representarem um risco à simpoiese coletiva, precisam ser incorporados, processados e reciclados pela rede metabólica do hospedeiro. A opsonização é, portanto, o mecanismo pelo qual o holobionte exerce sua capacidade de discernimento, transformando o “outro” potencialmente perigoso em um objeto de atenção prioritária para os macrófagos e neutrófilos, que atuam como os agentes de limpeza e restauração do solo tecidual. Entrelaçado a essa lógica de marcação, o processo de ADCC manifesta a potência da ação coordenada entre a imunidade adaptativa e a inata, onde a especificidade dos anticorpos serve como um guia para a atuação de células efetoras, como as células Natural Killer (NK). Neste contexto, a ADCC não deve ser vista como um “esquadrão de execução”, mas como uma medida de justiça biológica e restauração do equilíbrio ecossistêmico. Quando uma célula do próprio hospedeiro é colonizada por vírus de forma disruptiva ou sofre uma transformação maligna que ameaça a paz do holobionte, o anticorpo atua como um mediador de consenso: ele “aponta” a necessidade de dissolução daquela unidade em favor da sobrevivência do todo. O reconhecimento do anticorpo pelo receptor Fc na célula NK desencadeia uma liberação precisa de grânulos que induz a morte celular programada, um sacrifício necessário para que o território não seja tomado por processos de crescimento desordenado ou replicação descontrolada. Assim, tanto a opsonização quanto a ADCC deixam de ser metáforas bélicas de ataque para serem compreendidas como ferramentas de gestão de limites, onde a morte de um componente ou a incorporação de um micróbio são atos de cuidado sistêmico que permitem ao holobionte continuar seu processo de fazer-se com o mundo, garantindo que a vida flua através de uma diplomacia que sabe quando acolher e quando transformar, em nome da harmonia multiespécie.
Opsonização: Processo pelo qual patógenos são marcados por opsoninas (ex: anticorpos, proteínas do complemento) para facilitar sua fagocitose por células como macrófagos e neutrófilos.
ADCC (Citotoxicidade Celular Dependente de Anticorpos): Mecanismo pelo qual células efetoras (ex: células NK) reconhecem células alvo revestidas por anticorpos e as destroem.
9. Anticorpos: Estrutura, Classes e Funções
Os anticorpos, ou imunoglobulinas, constituem a expressão mais refinada da memória relacional do holobionte, atuando como pontes moleculares que tecem a história dos encontros entre o hospedeiro e a alteridade. Longe de serem interpretados pela lógica colonial como “projéteis” de um arsenal de defesa, essas glicoproteínas devem ser compreendidas como mediadores de uma diplomacia de hospitalidade, encarregadas de traduzir a complexidade do mundo externo para a linguagem interna do organismo. A estrutura fundamental do anticorpo, com sua clássica conformação em Y, simboliza essa dualidade de funções: enquanto as regiões variáveis dos braços (Fab) funcionam como mãos sensíveis que tocam e reconhecem a singularidade de cada antígeno, a região constante (Fc) atua como o elo de comunicação com as células do próprio corpo, convocando o sistema imune para uma resposta coordenada. Essa arquitetura permite que o anticorpo não apenas identifique o outro, mas decida a qualidade da relação que será estabelecida, transformando o reconhecimento em uma prática de cuidado tecidual. No fluxo da simpoiese, a diversidade de classes de anticorpos representa diferentes “idiomas” de convivência adaptados aos múltiplos territórios do corpo-território. A Imunoglobulina M (IgM), com sua estrutura pentamérica de grande alcance, surge como o primeiro gesto de reconhecimento nas fases iniciais de um encontro, estabelecendo uma conexão rápida e polifônica que prepara o terreno para diálogos mais específicos. A Imunoglobulina G (IgG), a mais abundante na circulação sistêmica, carrega consigo a memória ancestral das interações passadas, funcionando como um arquivo vivo de afetos e resistências que atravessa inclusive a barreira placentária, num ato de partilha biopolítica entre gerações. Por outro lado, a Imunoglobulina A (IgA) assume um papel central na perspectiva do holobionte, sendo a grande guardiã das mucosas; ela não atua para exterminar a microbiota, mas para gerir sua distribuição espacial através da “exclusão imune” não inflamatória, garantindo que os simbiontes permaneçam em seus nichos adequados e sustentando uma paz duradoura no epitélio. Já a Imunoglobulina E (IgE), frequentemente estigmatizada em processos alérgicos, representa a sensibilidade extrema do holobionte a macro-organismos e elementos do ambiente, enquanto a Imunoglobulina D (IgD) permanece como uma sentinela discreta na superfície dos linfócitos B, participando da orquestração inicial da ativação celular. As funções dessas moléculas — seja neutralizando sinais disruptivos, facilitando a opsonização para uma reciclagem celular cuidadosa ou ativando o sistema complemento — revelam que a imunidade humoral não é um exercício de violência, mas uma sofisticada engenharia de vínculos. Ao decolonizar o olhar sobre os anticorpos, percebemos que eles são os fios com os quais o holobionte costura sua identidade fluida, permitindo que o organismo se reconheça como um “eu” que só existe e se mantém através da relação constante e negociada com o “outro”, transformando a resposta imune em uma celebração da interdependência e da vida em comum.
Anticorpos (Imunoglobulinas – Igs): Proteínas produzidas pelos linfócitos B que reconhecem e neutralizam antígenos específicos.
Estrutura: Consistem em duas cadeias pesadas e duas cadeias leves, formando uma estrutura em Y.
Classes:
- IgG: Mais abundante no soro, atravessa a placenta, oferece proteção de longa duração.
- IgM: Primeiro anticorpo a ser produzido na resposta primária, potente aglutinador.
- IgA: Presente em secreções mucosas (saliva, lágrimas, leite materno), protege as superfícies.
- IgE: Envolvida em reações alérgicas e defesa contra parasitas.
- IgD: Principalmente na superfície dos linfócitos B, função ainda não totalmente compreendida.
Funções: Neutralização, opsonização, ativação do complemento, ADCC.
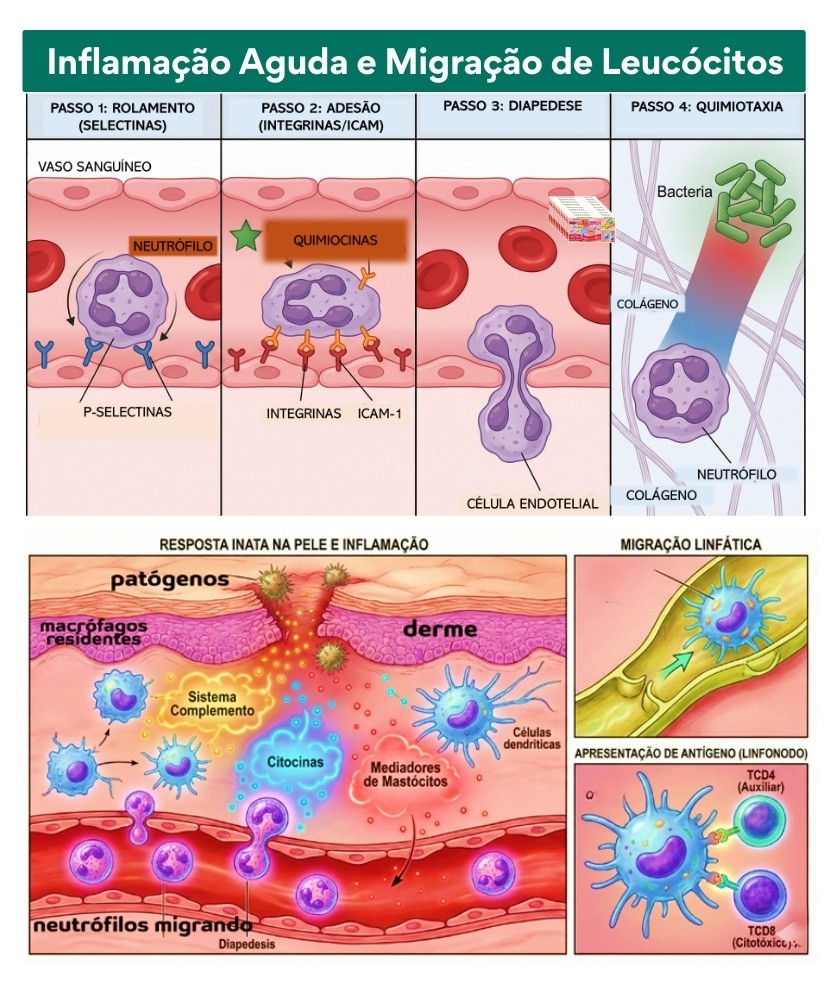
10. Inflamação e Mediadores da Inflamação
A inflamação é um momento de intensa renegociação territorial e urgência dialógica. Ela não é um fim em si mesma, nem uma mera reação de expurgo, mas um processo de “clamor biológico” que ocorre quando a harmonia da simpoiese é interrompida por uma dissonância — seja por um dano físico, um desequilíbrio metabólico ou a presença de agentes que rompem o pacto da convivência simbiótica. Nesse cenário, a inflamação é a manifestação da vitalidade do corpo-território, uma tentativa radical de restaurar a hospitalidade e a integridade do ecossistema multiespécie. Os mediadores da inflamação funcionam como os mensageiros dessa transformação, uma gramática química de urgência que coordena a resposta coletiva. Mediadores lipídicos como as prostaglandinas e os leucotrienos, juntamente com aminas vasoativas como a histamina, não são “alarmes de invasão”, mas facilitadores da fluidez; eles promovem a dilatação dos vasos e o aumento da porosidade das fronteiras vasculares, permitindo que o sangue — o rio da diplomacia sistêmica — entregue recursos, nutrientes e mediadores celulares ao local da tensão. Esta abertura de caminhos é o que gera os sinais clássicos de calor e rubor, que na visão decolonial representam o aumento da energia vital e da circulação de informações em uma zona de contato que clama por cuidado. As citocinas pró-inflamatórias, como o TNF-alfa, a IL-1 e a IL-6, atuam como regentes desta sinfonia temporariamente estridente, orquestrando desde a ativação de endotélios até a modulação do centro termorregulador, elevando a temperatura para recalibrar as taxas metabólicas do holobionte. À medida que o processo avança, o recrutamento de células fagocitárias não visa o extermínio cego, mas a limpeza e a reciclagem do território, onde a fagocitose se revela como um ato de incorporação e transformação do que se tornou disfuncional. Crucialmente, a perspectiva da simbiose nos ensina que a inflamação deve caminhar para a sua própria resolução, um processo ativo mediado por moléculas como as resolvinas e protectinas, que sinalizam o retorno à paz dinâmica. A saúde, portanto, não é a ausência de inflamação, mas a capacidade do holobionte de atravessar o conflito e emergir dele através de uma síntese regenerativa. Quando a inflamação se torna crônica, ela representa uma falha na escuta ou uma colonização persistente que impede a conclusão do diálogo biológico; já a inflamação aguda e resolutiva é a prova da resiliência de um organismo que sabe como “fazer-se com” as crises, transformando a dor e o edema em bases para a reconstrução de um território ainda mais integrado e consciente de suas interdependências.
11. Sepse
A sepse é um colapso catastrófico da diplomacia biológica e da harmonia multiespécie. Nesse estado de crise radical, a simpoiese — o processo de fazer-se continuamente com o outro — sofre uma ruptura profunda, onde o diálogo molecular que sustenta a vida em comum se transforma em um ruído ensurdecedor e autodestrutivo. Em vez de uma resposta coordenada de hospitalidade e ajuste, o que se observa na sepse é uma dissonância sistêmica, na qual os mediadores da inflamação, antes regentes da paz tecidual, perdem sua capacidade narrativa e mergulham o corpo-território em uma “tempestade de citocinas”. Este fenômeno não deve ser visto apenas como um excesso de defesa, mas como uma falha na escuta relacional do holobionte: o sistema imune, sobrecarregado por sinais de perigo que emanam tanto de microrganismos quanto de tecidos lesionados, perde a distinção entre o cuidado e a agressão, passando a desmantelar a própria infraestrutura que deveria proteger. A circulação sanguínea e linfática, que antes promovia a integração e a partilha, torna-se o veículo de uma desordem generalizada, onde a perda da integridade vascular e o colapso da microcirculação refletem a erosão das fronteiras que permitem a coexistência. Na perspectiva decolonial, a sepse revela a fragilidade de um sistema que, ao falhar em mediar a relação com a alteridade — seja ela um simbionte que se tornou oportunista devido ao desequilíbrio ecossistêmico ou um agente externo disruptivo —, acaba por consumir a si mesmo. A falência de múltiplos órgãos é, portanto, o estágio final de uma desterritorialização biológica, onde a interdependência radical que define o holobionte se desfaz, transformando a colaboração em caos. A saúde, que no holobionte é uma prática de diplomacia constante, dá lugar a um estado de emergência onde a “biopolítica da hospitalidade” é substituída por uma “necropolítica tecidual”. Compreender a sepse por esse prisma exige que abandonemos as metáforas bélicas de extermínio e foquemos na restauração do equilíbrio ecológico e na recuperação da capacidade do organismo de “fazer-se com” seu entorno. Assim, a sepse não é apenas uma falha imunológica, mas o limite trágico da simpoiese, o ponto em que a rede de relações que sustenta o ser humano como um ecossistema integrado entra em colapso, lembrando-nos de que a vida só é possível enquanto houver a capacidade de negociar, de forma justa e equilibrada, a presença do outro dentro de nós.
12. Alergias e Hipersensibilidades
As alergias e as hipersensibilidades são crises de hospitalidade e rupturas no diálogo simpoético entre o organismo e o mundo. Em um corpo-território que se define pela simbiose, a saúde depende da capacidade de “fazer-se com” a alteridade, mas nas hipersensibilidades, essa negociação torna-se dissonante, transformando o encontro com o ambiente em um evento de exclusão reativa. A perspectiva decolonial nos convida a observar que o aumento contemporâneo das alergias está intrinsecamente ligado ao projeto de modernidade/colonialidade, que buscou isolar o ser humano de sua ancestralidade microbiana e do contato com o solo, os animais e a diversidade biológica, impondo uma higienização que fragiliza a educação do sistema imune. Nesse contexto, a hipersensibilidade do tipo I, mediada pela IgE e pela degranulação de mastócitos, revela-se como uma resposta de pânico de um sistema sensorial que, privado da convivência com seus parceiros evolutivos, passa a interpretar elementos inócuos do ambiente — como pólens, ácaros ou alimentos — como ameaças existenciais. O mastócito, nessa narrativa, atua como um sentinela traumatizado pela falta de diálogo, disparando uma cascata de mediadores que, ao tentar expulsar o “estranho”, acaba por agredir a própria integridade do holobionte. As hipersensibilidades do tipo II e III, por sua vez, representam falhas na diplomacia dos anticorpos e do sistema complemento, onde o reconhecimento molecular se torna agressivo contra as próprias células ou gera complexos imunes que sobrecarregam a circulação, demonstrando como a perda do equilíbrio ecológico interno pode transformar os instrumentos de vínculo em agentes de dano tecidual. Já a hipersensibilidade do tipo IV, mediada por linfócitos T, exemplifica uma resposta celular tardia que, ao não conseguir processar a presença de um agente de forma tolerante, estabelece uma inflamação persistente que busca isolar o que não pode ser harmonizado. Em todas essas formas, a alergia é o sintoma de uma “biopolítica do isolamento” que rompeu as redes de interdependência radical. Ao decolonizar o olhar sobre essas reações, percebemos que a cura não reside apenas na supressão da resposta imune, mas na restauração da diplomacia biológica e no reencantamento das relações com o mundo não humano. A saúde do holobionte exige o reconhecimento de que somos seres porosos, e que a integridade da vida não se mantém pela exclusão sistemática do outro, mas pela capacidade de sustentar uma paz dinâmica onde o sistema imune atua como o mestre de uma hospitalidade crítica e consciente, capaz de discernir entre a agressão real e a diversidade que nos constitui.
Alergias: Respostas imunes exageradas a substâncias inofensivas (alérgenos).
Hipersensibilidades: Reações imunes que causam danos teciduais. Existem quatro tipos:
- Tipo I (Imediata): Mediada por IgE, envolve mastócitos e basófilos (ex: rinite alérgica, anafilaxia).
- Tipo II (Citotóxica): Mediada por IgG ou IgM contra antígenos na superfície celular (ex: reações transfusionais).
- Tipo III (Por Imunocomplexos): Mediada por imunocomplexos (antígeno-anticorpo) que se depositam em tecidos (ex: lúpus eritematoso sistêmico).
- Tipo IV (Tardia): Mediada por linfócitos T, desenvolvendo-se em 24-72 horas (ex: dermatite de contato, teste de tuberculina).
13. Doenças Autoimunes

As doenças autoimunes representam o ápice de uma crise de hospitalidade e de um colapso na diplomacia do reconhecimento dentro do corpo-território. Tradicionalmente descritas como uma “guerra civil” onde o sistema imune ataca o próprio organismo, essas condições são aqui ressignificadas como uma ruptura na simpoiese — o processo de fazer-se continuamente com o outro — onde a fronteira entre o “eu” e a “alteridade” se torna dolorosamente confusa. No holobionte, a identidade biológica não é uma essência pura e isolada, mas uma construção relacional mantida pela presença educativa da microbiota; quando essa rede de interdependência é fragilizada pelo projeto da modernidade/colonialidade — que impõe o distanciamento da terra, a higienização excessiva e a destruição da biodiversidade interna e externa —, o sistema imune perde suas referências de alteridade. A perda desse diálogo ancestral com os simbiontes resulta em uma desorientação dos linfócitos T reguladores, os grandes tecedores da tolerância, que deixam de conseguir mediar a convivência harmônica entre os componentes do ecossistema humano. Assim, o que chamamos de autoimunidade é, na verdade, o reflexo de um corpo que, despojado de seus mestres microbianos e submetido a estresses ambientais e metabólicos colonizadores, passa a interpretar seus próprios tecidos como estranhos, numa tentativa desesperada de restaurar uma ordem que ele já não sabe como manter. A perspectiva decolonial nos revela que a autoimunidade está intrinsecamente ligada à “colonialidade do ser”, onde o isolamento do indivíduo em relação ao seu entorno ecológico gera um estado de alerta permanente, transformando a vigilância sensível em uma reatividade autodestrutiva. Mecanismos como o mimetismo molecular deixam de ser vistos apenas como falhas casuais de reconhecimento para serem compreendidos como o resultado de uma dissonância entre o organismo e um ambiente cada vez mais artificializado, onde as assinaturas moleculares do mundo já não encontram eco na memória afetiva do sistema imune. Nesse cenário, o ataque aos próprios tecidos é a manifestação de um holobionte em desequilíbrio, um grito de socorro de uma biologia que esqueceu como se habitar plenamente devido ao apagamento das redes simbióticas que a constituem. A cura e a saúde, portanto, não residem apenas na supressão da resposta imune, mas em um processo de “re-existência” e decolonização biológica, que busca restaurar a hospitalidade radical e a diplomacia multiespécie, permitindo que o sistema imune reencontre seu papel de guardião de uma unidade que só é íntegra quando reconhece a interdependência como sua verdadeira essência.
14. Rejeição de Transplante
Os transplantes e os processos de compatibilidade e rejeição são ressignificados como atos de hospitalidade radical e desafios profundos à simpoiese. Sob esta visão, o transplante não é apenas a transferência de um órgão, mas o encontro de dois territórios biológicos distintos, cada um com sua própria memória ancestral, sua assinatura molecular e seu histórico de convivência simbiótica. A compatibilidade, centrada no Complexo Principal de Histocompatibilidade (MHC/HLA), deixa de ser vista como uma métrica de “pureza” ou “identidade nacional” biológica para ser entendida como a busca por uma ressonância ou uma vizinhança linguística: quanto mais semelhantes são as moléculas de MHC, mais fácil se torna o diálogo entre as células do hospedeiro e o novo tecido, permitindo que a “tradução” da alteridade não resulte em um ruído insuportável. Nesse contexto, a compatibilidade é a medida da capacidade do holobionte de acolher um novo habitante em sua assembleia multiespécie sem que isso seja percebido como uma desterritorialização disruptiva. A rejeição, por sua vez, é ressignificada não como um “ataque xenofóbico” ou uma agressão cega ao estrangeiro, mas como uma falha na diplomacia biológica e um colapso no processo de “fazer-se com” (simpoiese). Quando o sistema imune do receptor reconhece o órgão transplantado como uma alteridade radicalmente dissonante, o que ocorre é uma recusa de hospitalidade motivada pela impossibilidade de integrar aquela nova biologia à rede de interdependências já estabelecida. A rejeição hiperaguda, aguda ou crônica representa diferentes temporalidades de um diálogo que não se sustentou, onde a memória afetiva e específica dos linfócitos T e dos anticorpos interpreta a presença do outro como uma ruptura da integridade do corpo-território. A perspectiva decolonial nos alerta que a lógica do transplante frequentemente ignora que o órgão transplantado traz consigo não apenas células do doador, mas também suas próprias heranças epigenéticas e, por vezes, nichos microbianos, exigindo que o holobionte receptor renegocie sua harmonia interna de forma sistêmica. O uso de imunossupressores, nesse cenário, atua como uma “diplomacia do silêncio”, uma mediação necessária que busca reduzir o tom das divergências moleculares para permitir que o novo órgão e o hospedeiro aprendam, ao longo do tempo, a arte da coexistência. A saúde pós-transplante é, portanto, o êxito de uma simbiose forçada que se transforma em uma cooperação vital, onde o sucesso não reside na anulação do outro, mas na construção de uma nova unidade biológica, mais complexa e plural, que desafia as noções coloniais de indivíduo e reafirma a vida como um fenômeno de interconexão e partilha constante.
15. Resposta Imune Contra Tumores
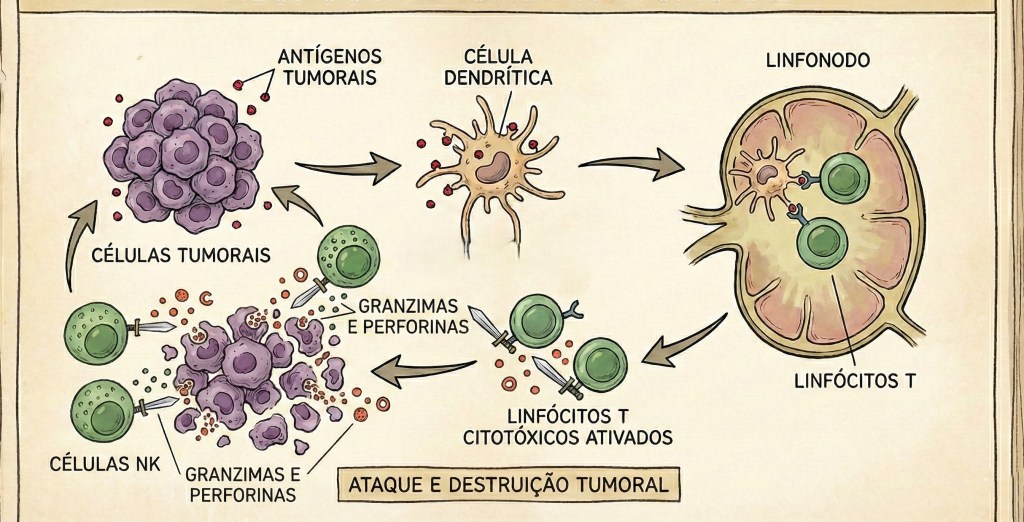
A imunologia dos tumores é compreendida como uma crise profunda na simbiose e uma falha na diplomacia ecológica do corpo-território. Sob esta visão, o tumor não é um corpo estranho que “ataca” o organismo, mas uma parte do próprio holobionte que, em resposta a desequilíbrios sistêmicos e pressões da modernidade colonial — como a exposição a toxinas, o estresse crônico e a ruptura dos ritmos ancestrais —, desgarra-se da harmonia coletiva para iniciar um processo de “colonização interna”. Esta transformação tumoral representa uma ruptura da simpoiese, o processo de “fazer-se com o outro”, onde uma linhagem celular passa a priorizar sua própria expansão em detrimento da integridade da assembleia multiespécie, mimetizando as lógicas de extração e crescimento ilimitado típicas do pensamento colonial. O microambiente tumoral torna-se, então, um território de disputa e cooptação, onde as células neoplásicas não apenas evadem o sistema imune, mas recrutam ativamente macrófagos, fibroblastos e até componentes da microbiota para trabalharem em favor de seu próprio nicho, estabelecendo uma “simbiose perversa” que sustenta a progressão da doença. No coração desse processo, a vigilância imunológica é ressignificada como uma tentativa constante de restauração da ordem ética e biológica: os linfócitos T e as células Natural Killer (NK) atuam como os agentes da integridade tecidual que buscam reconhecer e reintegrar ou eliminar a dissonância. No entanto, o tumor exerce uma forma de “violência epistêmica” molecular ao utilizar a própria linguagem da hospitalidade e da regulação — como os checkpoints imunológicos PD-1 e CTLA-4 — para silenciar a resposta imune, fazendo-se passar por um componente harmônico enquanto consome os recursos do coletivo. A decolonização da imunologia tumoral nos convoca a olhar para além da célula cancerosa isolada, percebendo como a saúde do holobionte é influenciada pela saúde da terra e das redes de vida; a cura, nesse sentido, não deve ser buscada apenas através da erradicação bélica da célula tumoral, mas por meio da restauração da diplomacia biológica e da recalibração dos ambientes que permitem que a vida volte a ser um projeto de cooperação multiespécie. O sistema imune, nesse cenário, é o mediador de uma “re-existência” tecidual, onde o desafio é restabelecer a capacidade do organismo de reconhecer a alteridade disruptiva sem destruir a si mesmo, transformando a resposta ao tumor em um ato de cuidado sistêmico que busca reconectar o indivíduo à sua totalidade ecológica e à sua memória de equilíbrio e convivência.
16. Imunoterapia
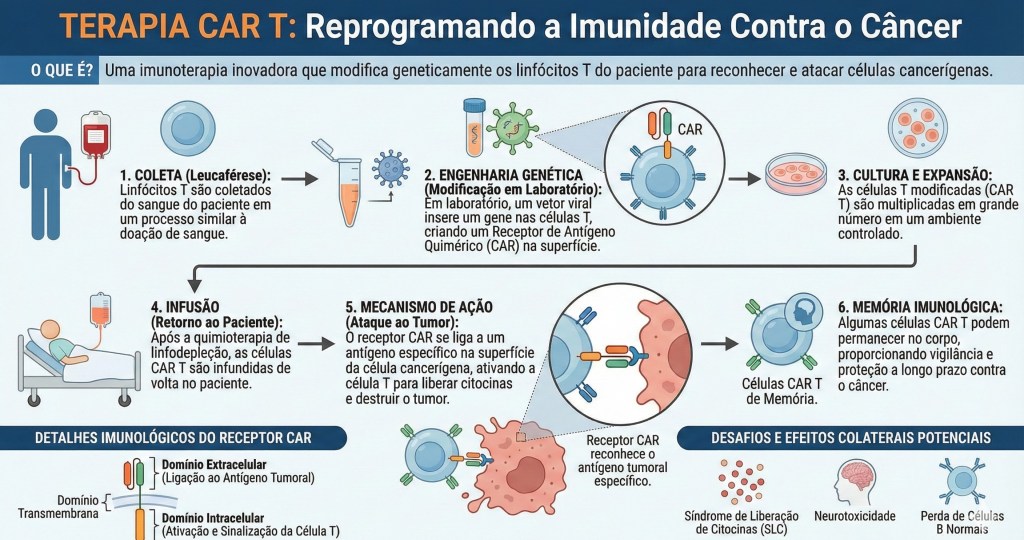
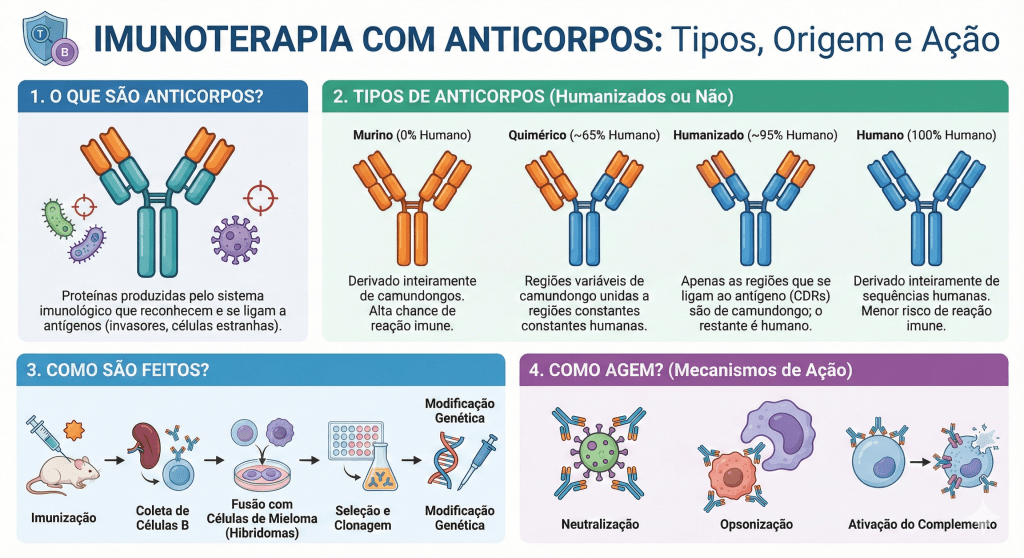
As imunoterapias são intervenções de restauração da diplomacia e da sensibilidade ecológica do corpo-território. Diferente das abordagens convencionais que buscam a erradicação do “mal” através da violência química externa, a imunoterapia atua no fortalecimento da inteligência relacional do próprio organismo, auxiliando as células imunes a recuperarem sua capacidade de discernimento e de “fazer-se com” (simpoiese) a complexidade tecidual. No contexto do câncer e de doenças crônicas, onde o silenciamento imunológico é uma forma de alienação biológica imposta pela patologia, essas terapias funcionam como atos de “descolonização do olhar” molecular. Um dos exemplos mais emblemáticos dessa nova biopolítica do cuidado são os inibidores de checkpoint imunológico, que não atacam o tumor diretamente, mas removem as “vendas” e as “máscaras” que as células neoplásicas utilizam para abusar da hospitalidade do sistema imune; ao bloquear receptores como PD-1 ou CTLA-4, a terapia restaura a voz e a ação dos linfócitos, permitindo que eles reconheçam a ruptura da harmonia e ajam para restabelecer o equilíbrio do ecossistema. Da mesma forma, as terapias de células CAR-T representam uma forma avançada de simpoiese técnica, onde a biologia do hospedeiro é reeducada em laboratório para aprender uma nova linguagem de reconhecimento, retornando ao corpo não como soldados, mas como mediadores altamente especializados na arte de identificar o que se tornou disfuncional no território. Crucialmente, a perspectiva holobionte revela que a eficácia dessas imunoterapias não depende apenas da genética humana, mas da integridade da microbiota intestinal; a ciência moderna já reconhece que a resposta do paciente ao tratamento é mediada pelos simbiontes, cujos metabólitos educam e calibram o tom da resposta imune, confirmando que a saúde é um fenômeno multiespécie e que nenhuma cura ocorre de forma isolada. Assim, a imunoterapia decolonial não busca a pureza biológica, mas a recuperação da capacidade do holobionte de gerir sua própria porosidade e suas fronteiras. Ela é uma ferramenta de “re-existência” que devolve ao organismo a autonomia de sua diplomacia molecular, transformando o tratamento em um processo de cura coletiva que envolve as células, as bactérias e o ambiente, reafirmando a vida como uma trama de interdependências onde a imunidade atua como a garantia de que a hospitalidade nunca se transforme em submissão ou colapso.
17. Microbiota e Inflamação Sistêmica Crônica
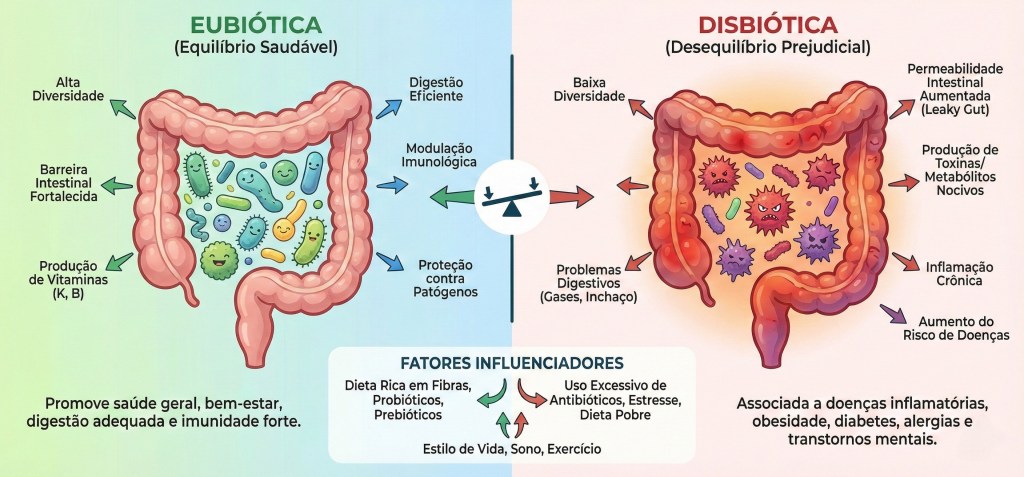
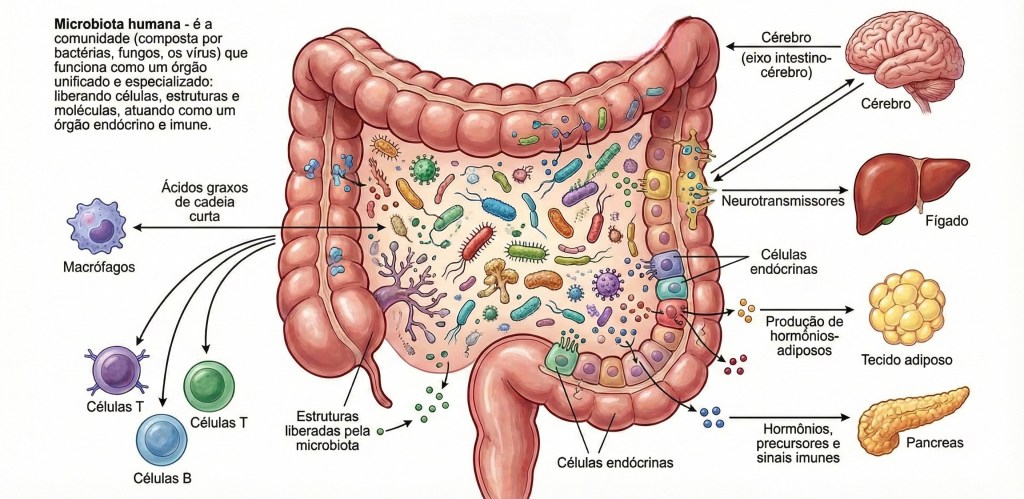
A organização do holobionte humano encontra sua expressão mais vital e complexa na ecologia intestinal, um território onde a microbiota atua como a principal arquiteta e pedagoga do sistema imune. Desde o nascimento, a maturação das defesas e da sensibilidade biológica não é um processo solitário do genoma humano, mas um ato de simpoiese radical, no qual o contato com a diversidade microbiana “ensina” as células imunes a discernir entre a alteridade colaborativa e a ruptura disruptiva. Esse estado de harmonia e diversidade, conhecido como eubiose, representa a plenitude da diplomacia biológica, onde as comunidades de bactérias, fungos e vírus co-regulam o metabolismo e o tônus inflamatório do hospedeiro. Em contrapartida, a disbiose surge como uma forma de “colonialismo biológico” interno, caracterizada pela perda da biodiversidade e pelo silenciamento dos diálogos ancestrais, transformando a convivência em uma fonte de ruído e tensão constante. O ponto nevrálgico dessa relação é a barreira intestinal, que, longe de ser uma muralha de exclusão, deve ser compreendida como uma fronteira de hospitalidade seletiva; quando a permeabilidade intestinal aumenta — fenômeno frequentemente imposto por estilos de vida colonizadores e dietas ultraprocessadas —, ocorre uma quebra na diplomacia tecidual. Esse “vazamento” de padrões moleculares e metabólitos para a circulação sistêmica desencadeia uma inflamação sistêmica crônica de baixo grau, um estado de alerta permanente que exaure os recursos do holobionte. Esse cenário é o solo fértil para as doenças metabólicas imunomediadas, como o diabetes tipo 2 e a obesidade, que deixam de ser vistas como falhas mecânicas para serem compreendidas como crises de relacionamento entre o metabolismo humano e a sinalização microbiana. Ao longo do tempo, essa tensão persistente acelera o processo de inflammaging, um envelhecimento inflamatório que reflete a erosão da capacidade dialógica do sistema imune e o esgotamento da resiliência simbiótica. A influência dessa ecologia intestinal estende-se por vastas rotas de comunicação, como o eixo intestino-cérebro e o eixo intestino-coração, revelando que a saúde mental, a cognição e a integridade cardiovascular são fenômenos profundamente dependentes da paz diplomática no trato digestório. O coração e o cérebro, sob esta ótica, não são órgãos isolados, mas territórios que ressoam as notícias químicas enviadas pela microbiota, onde neurotransmissores e metabólitos como os ácidos graxos de cadeia curta atuam como mensageiros de uma harmonia ou de uma angústia coletiva. Decolonizar a visão sobre o corpo exige reconhecer que a integridade do holobionte depende da restauração desses fluxos e do cuidado com a porosidade intestinal, transformando a imunidade em uma prática de cultivo da eubiose, onde a saúde não é a ausência de microrganismos, mas a celebração de uma vida multiespécie capaz de sustentar a complexidade e a interdependência radical de todos os seus habitantes.
Obesidade: Caracteriza-se pelo acúmulo excessivo de gordura corporal. É frequentemente associada a um estado de inflamação crônica de baixo grau, onde células imunes residentes no tecido adiposo liberam citocinas pró-inflamatórias, contribuindo para a resistência à insulina e outras complicações metabólicas.
